
文/張瑞宗、廖竣、林威任、陳若尹、曾晴賢
.png) 外來異形魚在臺灣
外來異形魚在臺灣
臺灣的觀賞水族產業始於1960年代,出口貿易的產業型態多半是捕撈海水魚隻販售至其他國家為主,國內水族市場的流通商品則多以錦鯉、金魚為大宗。1980年代後,由於經濟起飛所帶動的消費行為,臺灣的觀賞水族業開始蓬勃發展,進而陸續進口他國水生生物,從非洲的麗魚科(Cichlidae)魚類至南美洲亞馬遜流域的脂鯉科(
Characidae)魚類等皆是極具高觀賞價值的熱門物種。在水族產業中,有些物種的引進是根據該物種之功能性,如現在水族市場泛稱的異形魚多半具有撿食水族缸中殘餌、清除缸壁附著藻類等益處而引進作為工具魚之用。坊間常見的異形魚有不少種類皆原產於中、南美洲,而著名的亞馬遜河流域,其流域面積廣闊且地形多變,更是演化與孕育出各式各樣異形魚的重要種源地之一。目前已知流通於水族市場的異形魚,在分類上屬於條鰭魚綱(Actinopterygii)、新鰭魚亞綱(Neopterygii)、鯰形目(Siluriformes)、甲鯰科(Loricariidae)魚類,臺灣的原生種淡水魚類並未包含甲鯰科魚類成員,然而外來異形魚的入侵卻是早已充斥著臺灣各大小溪流、湖泊、埤塘、水庫等淡水水域環境中,成為眾所皆知的外來種課題。
目前已知入侵臺灣的外來種異形有兩種,一為早在約莫1970年代即被引進臺灣,俗稱「垃圾魚」、「清道夫」又或是「琵琶鼠」的翼甲鯰魚類,因其對臺灣淡水域的適應良好,再加上此物種對汙染與低溶氧的耐受性較高,全身披覆硬質骨板不易遭掠食者掠食,經過多年來的繁衍與快速入侵,至今翼甲鯰已入侵至臺灣各大小淡水水域環境中,成為難以完全根除的外來入侵魚種。根據臺灣魚類資料庫的紀錄,引進臺灣的翼甲鯰屬(
Pterygoplichthys)魚類應該至少有野翼甲鯰(
Pterygoplichthys disjunctivus)與豹紋翼甲鯰(
P. pardalis)兩物種,而野翼甲鯰的最大體長可達70公分,而豹紋翼甲鯰的最大體長至少也有40公分以上,但有研究證實翼甲鯰屬這類群的魚類在不同種間會互相雜交,因此當今入侵臺灣野外的翼甲鯰多半為野翼甲鯰與豹紋翼甲鯰的雜交個體,本文則先以雜交翼甲鯰稱之(圖1)。
 圖1. 已入侵全臺大小水域的雜交翼甲鯰(Pterygoplichthys sp.)
圖1. 已入侵全臺大小水域的雜交翼甲鯰(Pterygoplichthys sp.)
另一入侵臺灣的異形魚則是本年初(2021年)於臺中市霧峰區的阿罩霧圳發現俗稱為「鬍子異形」的已入侵鈎鯰屬(
Ancistrus)魚類,其與翼甲鯰魚類於分類系統中同屬於甲鯰科下的魚類。鈎鯰屬魚類目前已有明確被鑑定出的物種至少有68種,在分類上多以鼻部前緣在成長時發育出的皮質觸鬚型態作為鑑種的分類依據,然而在種間的型態上因具有高度的相似度,故截至目前尚存有許多未被描述的新物種,倘若包含尚未描述的隱匿物種,目前初估鈎鯰屬的物種至少有90個物種以上(Alves
et al., 2006)。有些研究為了釐清鈎鯰屬魚類的性別決定機制,發現鈎鯰屬的各物種至少就包含了5種性別決定機制,分別為ZZ/ZW、XX/XY、XX/XO、XX/XY1Y2與Z1Z1Z2Z2/Z1Z2W1W2,亦記錄到鈎鯰屬中有些不同的物種之間可進行雜交並產下子代(Alves et al., 2006)。鈎鯰屬魚類的細胞核型染色體經統計後發現不同的物種其染色體總對數的變異最少有17對,多則達到26對,顯示其染色體重組的核型演化正發生在鈎鯰屬的各物種間,代表鈎鯰屬的魚類保有相當大的多樣性(Mariotto
et al., 2004;Alves
et al., 2006;Oliveira
et al., 2007)。入侵臺灣的鈎鯰屬魚類,不同的個體其體色與型態差異很大,甚至還發現了在水族市場流通經過選育的黃化個體,故入侵臺灣的鈎鯰屬魚應屬於哪一物種,仍需利用分子生物鑑定方式再進一步確認,方能有所頭緒。這尚未確認物種的入侵鈎鯰屬魚類,在後續的文章中則先以鬍子異形撰述(圖2、圖3)。
 圖2. 2021年發現的新興已入侵鬍子異形(Ancistrus sp.)個體
圖2. 2021年發現的新興已入侵鬍子異形(Ancistrus sp.)個體
 圖3. 鬍子異形(Ancistrus sp.)入侵場域所發現的黃化鬍子異形個體
圖3. 鬍子異形(Ancistrus sp.)入侵場域所發現的黃化鬍子異形個體
雜交異甲鯰與鬍子異形比一比
入侵臺灣的雜交翼甲鯰與鬍子異形雖同屬於甲鯰科的魚類,因此在外觀上有許多相似的特徵,如兩物種的魚體皆披覆硬質骨板,然而在分類上,兩物種一為翼甲鯰屬一為鈎鯰屬,不同屬之間物種仍舊可發現牠們在外觀上的些微差異,為了能讓讀者更了解兩入侵物種間的差異並更加認識牠們。我們將兩物種的差異整理出一簡單的表格,提供給大家參考(表1)。
表1. 雜交翼甲鯰與鬍子異形鑑別特徵對照表

阿罩霧圳入侵鬍子異形的發現
烏溪起源於北合歡山,流經臺中市與彰化縣,烏溪以北為臺中市由上游至下游分別流經霧峰、烏日、大肚與龍井等地;以南為彰化縣。霧峰地區位於烏溪中上游,此段河道相較於下游而言,河道較小且坡度較為陡急(馬,2008);霧峰地區亦位處於車籠埔斷層與隘寮斷層帶的交界處,經長時間斷層帶運動下,將附近區域的地層大致分為2種地層,厚度約6~8公尺的砂礫石層與岩盤,其中岩盤的性質以泥質砂岩與頁岩為主,且依據ISRM分類屬於軟弱岩盤(李等, 2019)。早期霧峰地區常面臨水患等問題,日治時期臺灣總督府針對烏溪的治水事業,歷時約40餘年,前後進行水勢與地質調查、治水設施的興建與修復、鄰近區域埤圳的串聯等規劃及執行,同時林獻堂也積極參與建設與修復;如日治初期合併丁臺圳、南勢圳並改稱為阿罩霧圳(圖4),由林獻堂擔任管理人,爾後在林獻堂多次針對護岸進行提議、維護與興建等事宜,使得阿罩霧圳的灌溉水利設施得以健全,建構起霧峰地區農業發展的根基。如今,臺中市的霧峰地區,是臺灣農產品的重點培育區域之一,而幕後的功臣,便是流遍霧峰區的阿罩霧圳,其目前由行政院農業委員會水利署南投管理處管轄,至今仍然為霧峰地區重要的灌溉水利設施。
 圖4. 阿罩霧圳自烏溪將水源引入並流經臺中市霧峰區
圖4. 阿罩霧圳自烏溪將水源引入並流經臺中市霧峰區
隨著時代的進步,網路加速了資訊的更新,早於去年年底(2020年)即有民眾拍攝到在野溪環境中捕獲鬍子異形的影片於網路流傳,今年年初(2021年)亦有民眾團隊拍攝捕捉到鬍子異形的影片於網路宣導勿隨意棄養外來種之議題。為了釐清網路消息正確與否?以及鬍子異形可能的入侵範圍?本團隊前往位在臺中市霧峰區的阿罩霧圳排水系統進行調查與記錄。阿罩霧圳於第一分水堰後分流為阿罩霧排水第一幹線與阿罩霧排水第二幹線,而在這兩排水幹線皆有發現鬍子異形入侵的個體。因阿罩霧排水第二幹線有一段流經農業試驗所轄區內,因此無法隨意進入農業試驗所場域內進行調查並確認鬍子異形的入侵狀況;阿罩霧排水第一幹線則流經農業試驗所正門口,經匯流丁台圳後成為頭汴坑溪,最後匯入大里溪後再流入烏溪主流。經初步調查後發現,鬍子異形尚未入侵到下游的頭汴坑溪段,目前較明確的入侵的範圍為仍以阿罩霧圳第一幹線為主要範圍,且在調查過程中亦陸續捕獲鬍子異形之幼魚,證明鬍子異形已適應臺灣野溪環境並成功繁衍(圖5)。
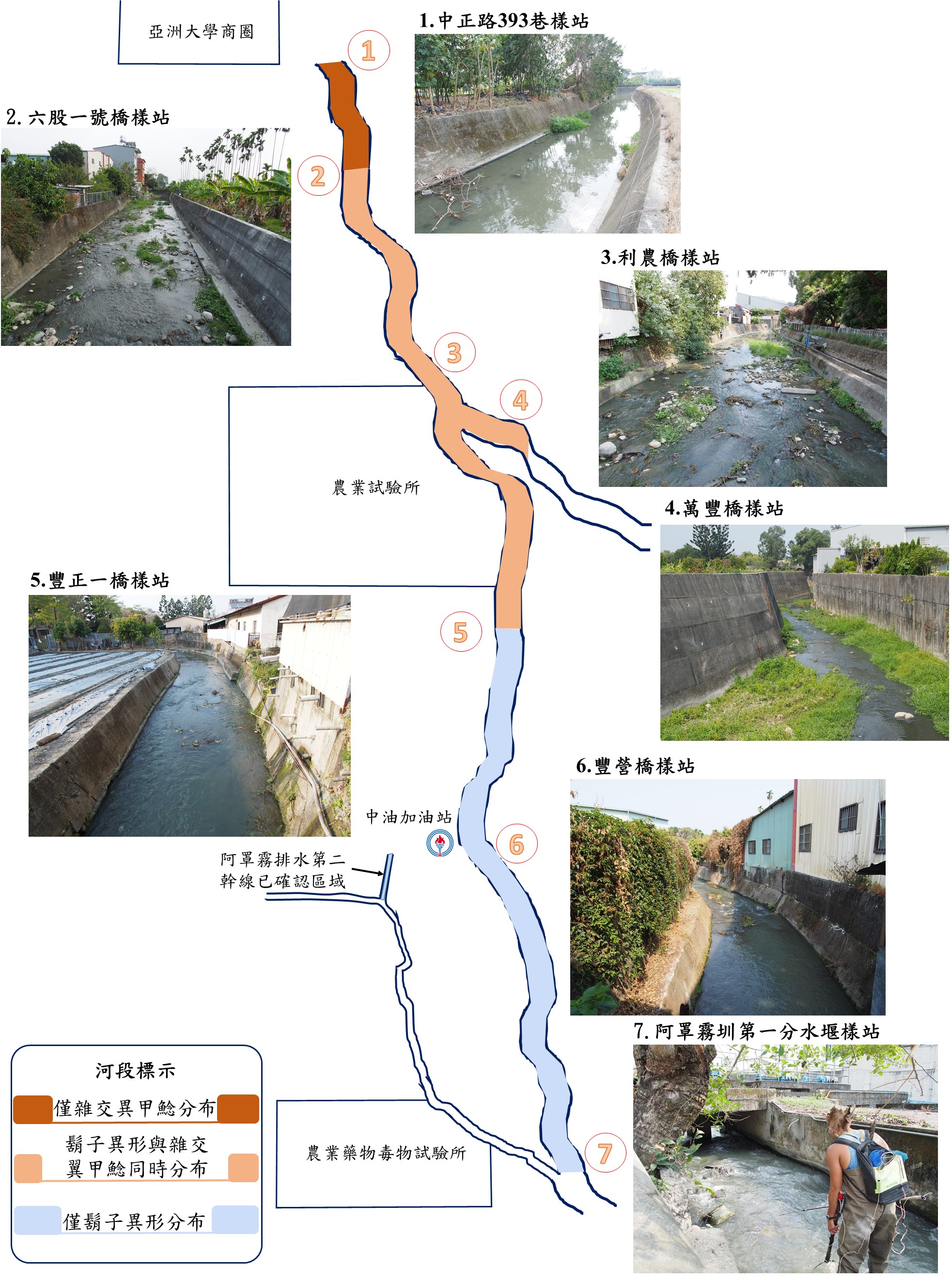 圖5. 阿罩霧排水第一幹線各調查樣站與鬍子異形、雜交翼甲鯰入侵分布概況
圖5. 阿罩霧排水第一幹線各調查樣站與鬍子異形、雜交翼甲鯰入侵分布概況
入侵異形魚住的不一樣!
在阿罩霧排水第一幹線可發現入侵的雜交翼甲鯰與鬍子異形兩大物種,根據調查的結果顯示,鬍子異形入侵的分布界線,大致以阿罩霧排水第一幹線上游的阿罩霧圳第一分水堰樣站(圖6)至下游的六股一號橋樣站(圖7)為其目前的入侵分布範圍,六股一號橋樣站往下游則暫無捕獲記錄,然而其分布範圍是否會往上游烏溪方向入侵或是往下游河段持續擴散是亟需要持續進行追蹤的;雜交翼甲鯰的分布則是以豐正一橋樣站(圖8)為界,此樣站往上游暫無捕獲記錄,豐正一橋樣站往下游則皆有雜交翼甲鯰的捕獲記錄。彙整兩甲鯰科魚類入侵的分布資訊後,發現位於上游豐正一橋樣站至下游六股一號橋樣站之間的河段,可同時發現鬍子異形與雜交翼甲鯰兩入侵物種, 且發現鬍子異形在阿罩霧排水第一幹線越往上游的樣站,採捕到的個體數量越多;反之,雜交翼甲鯰的入侵個體則是越往阿罩霧排水第一幹線的下游樣站捕獲的數量越多。由此可知此兩入侵甲鯰科魚類在阿罩霧排水第一幹線的入侵趨勢,越往上游的河段,鬍子異形較為優勢,越往下游則是雜交翼甲鯰較為優勢(圖5、圖9)。
 圖6. 阿罩霧圳第一分水堰將烏溪引入的水源分流給阿罩霧排水第一幹線與阿罩霧排水第二幹線
圖6. 阿罩霧圳第一分水堰將烏溪引入的水源分流給阿罩霧排水第一幹線與阿罩霧排水第二幹線
 圖7. 阿罩霧排水第一幹線的六股一號橋下游為泥質底流速較為緩慢的棲地類型
圖7. 阿罩霧排水第一幹線的六股一號橋下游為泥質底流速較為緩慢的棲地類型
 圖8. 阿罩霧排水第一幹線的豐正一橋樣站可見民宅的汙水排水管
圖8. 阿罩霧排水第一幹線的豐正一橋樣站可見民宅的汙水排水管
 圖9. 阿罩霧排水第一幹線各樣站捕捉到入侵甲鯰科兩物種的比例圖
圖9. 阿罩霧排水第一幹線各樣站捕捉到入侵甲鯰科兩物種的比例圖
翻閱相關文獻後,發現鬍子異形的原生棲息環境是水流較為湍急,且水質較乾淨的水域。阿罩霧排水第一幹線流經霧峰區的農田、工廠與民宅,隨著越往中、下游,河段累積的農業、工廠與民宅所產生的汙染物質越來越多,故越往下游其水質條件越不利於對汙染耐受性較低的鬍子異形,也因此取而代之的是對環境汙染耐受性較高的雜交異甲鯰。另外在鬍子異形對棲息地的選擇,喜愛具水流的石縫或是有良好遮蔽效果的洞穴為主,恰巧阿罩霧排水第一幹線的流況,越往下游流速相對緩慢且泥質底的緩流區域亦較多,故鬍子異形並不會利用此類型的棲地環境;然而雜交翼甲鯰雖然有時可在較有流速的水域中發現,但其偏好利用的棲地環境仍是流速相對緩慢的洞穴中或場域,故越往下游自然更有利於入侵的雜交翼甲鯰的族群。
鬍子異形的特殊繁衍行為
鬍子異形喜愛有水流且棲居於水中倒木或是岩石所堆疊形成的天然遮蔽處下,並建立屬於自己的領域範圍。繁殖季到來時,雄魚會在交配前先尋找合適的產卵區,接著清理產卵區周圍直到雌魚願意靠近產卵,雌魚在選擇雄魚時,已護幼的雄魚亦較有機會成為交配繁衍的對象。當完成交配後,雄魚會負責保護授精卵,並照護卵粒與剛孵化出的年幼個體,使其免於受到掠食者掠食,此護幼行為會持續至受精卵孵化後約10天左右結束,護幼期間雄魚會積極地抵禦任何可能的威脅,藉此提高稚魚的存活率(Sabaj
et al., 1999)。為了更進一步釐清已在臺灣入侵的鬍子異形的繁殖季節,本團隊自本年度(2021年)3月與4月所捕獲的入侵鬍子異形分別抽樣解剖127尾與108尾個體,觀察其生殖腺成熟度,希望藉此資訊了解其繁殖狀況。目前初步確認於阿罩霧排水第一幹線捕獲的鬍子異形已有性成熟之雌雄個體,加上於阿罩霧排水第一幹線的固定樣站進行監測與移除作業時所發現小型年幼個體之紀錄,可再次確認鬍子異形已能適應臺灣淡水環境並達到性成熟,更進一步地能成功繁衍後代。進一步統計後發現3月份的解剖個體中有33%的個體達性成熟,4月份的解剖個體中則有高達38%的個體達性成熟,此結果亦看出自3月起,入侵阿罩霧排水第一幹線的鬍子異形有逐漸進入繁殖高峰的趨勢(圖10)。根據雌性的鬍子異形,經細算卵巢之孕卵數後發現成熟性腺平均至少可計數到91粒卵粒,體型最大的性成熟雌性鬍子異形體長為8.6cm,孕卵數可達為225粒;體型最小的性成熟雌性個體體長僅為5.2cm,孕卵數有48粒,亦有發現體長為5.4cm的雌性性成熟個體,其孕卵數高達118粒。相近體長的雌性性成熟個體,雖孕卵數因生活環境及攝食狀況有所落差,但孕卵數可高達225粒的鬍子異形再加上護幼行為增加了此物種快速入侵與擴散至臺灣野溪的可能。然而目前對新興入侵臺灣野溪的鬍子異形其生殖習性,如:生殖期長短、頻率與孵化率等細節皆尚未有明確的研究,因此除了對此物種持續地進行監測與移除工作外,對其繁衍行為的釐清更是刻不容緩。
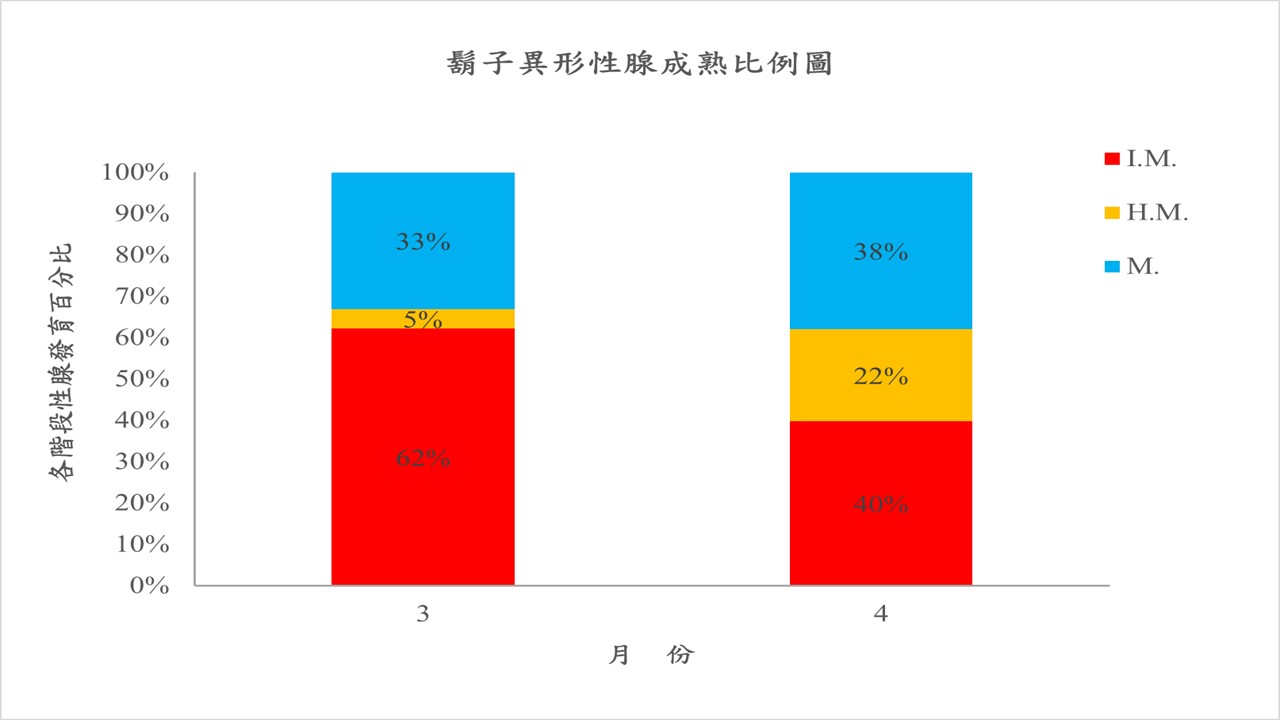 圖10. 鬍子異形性腺成熟比例圖(M.:表性腺成熟、H.M.:表性腺半成熟、I.M.:表未成熟性腺)
圖10. 鬍子異形性腺成熟比例圖(M.:表性腺成熟、H.M.:表性腺半成熟、I.M.:表未成熟性腺)
 圖11. 性腺成熟的雄性鬍子異形可見一對白色的精巢
圖11. 性腺成熟的雄性鬍子異形可見一對白色的精巢
 圖12. 性腺成熟的雌性鬍子異形可見飽滿的卵巢
圖12. 性腺成熟的雌性鬍子異形可見飽滿的卵巢
 圖13. 顯微鏡下的鬍子異形卵粒外觀
圖13. 顯微鏡下的鬍子異形卵粒外觀
結語
相較於過往,當今已有不少民眾對已入侵的外來水生魚類議題有一定程度的了解,對於甲鯰科魚類的雜交翼甲鯰更是有不少論及入侵臺灣的相關專題報導。同為甲鯰科魚類的鬍子異形,卻是本年度(2021年)才剛發現的新興外來入侵物種,針對其對臺灣淡水原生物種的衝擊除了過往已知甲鯰科因其特殊的覓食行為,會使得許多原生魚類的受精卵被攝食外,另外推測亦有可能對臺灣原生淡水魚類在棲息空間上有所排擠,然而其對社會經濟的損失是否另有其他嚴重的威脅,目前尚不明朗。在掌握目前鬍子異形已入侵的場域目前上侷限在阿罩霧排水幹線的我們,除了基礎的定期監測與移除工作外,此物種是否會藉由引水灌溉渠道逐漸往上、下游水域擴散,這才是我們目前最擔憂的狀況。外來入侵種的入侵如同新冠肺炎的疫情般,當發現的入侵案例的初期就得當機立斷,即刻進行防堵其擴散與針對入侵個體的移除作業,在其仍為星星之火之時若不立即撲滅,即可能導致不可收拾的嚴重後果。相信有不少關心臺灣淡水生態的朋友都不希望見到鬍子異形的入侵成為外來種雜交翼甲鯰第二,為了避免此窘境,除了中央政府的移除政策外,結合在地民眾或在地社區的力量一同來防堵,或許更可提高不少抑制入侵族群的效能唷!
1.李宗庭、李偉哲、莊育蓁、黃俊仁、洪炳榮。2019。人工湖設計經驗談-以烏溪鳥嘴潭人工湖為例。中興工程。144:61-69。
2.吳筱萍。2002。高屏溪琵琶鼠族群特徵、生殖週期與食性之研究。國立高雄師範大學。碩士論文。64頁。
3.馬鉅強。2008。烏溪治水事業之研究—以《灌園先生日記》為中心。臺灣文獻季刊。58(1):245-288。
4.劉素芬。2007。日治時期霧峰水利組織與地域社會的演變。中央大學人文學報。31:145-180。
5.Alves A.L., Oliveira C., Nirchio M., Granado A. and Foresti F., 2006, Karyotypic relationships among the tribes of Hypostominae (Siluriformes: Loricariidae) with description of XO sex chromosome system in a Neotropical fish species., Genetica, 128: 1–9.
6.Mariotto S., Artoni R.F. and Miyazawa C.S., 2004, Occurence of sexual chromosome, of the type ZZ/ZW, in Ancistrus cf. dubius (Loricariidae, Ancistrinae) of the Paraguay River Basin, Mato Grosso, Brazil., Caryologia, 57: 327–331.
7.Sabaj M.H., J.W. Armbruster and L. M. Page., 1999, Spawning in Ancistrus (Siluriformes: Loricariidae) with comments on the evolution of snout tentacles as a novel reproductive strategy: larval mimicry., Ichthyological Exploration of Freshwaters., 10(3): 217-229.
8.Oliveira R.R., Feldberg E., Anjos M.B. and Zuanon J., 2007, Karyotype characterization and ZZ/ZW sex chromosome heteromorphism in two species of the catfish genus Ancistrus Kner, 1854 (Siluriformes: Loricariidae) from the Amazon basin., Neotropical Ichthyology, 5: 301–306.
張瑞宗
清華大學生物資訊與結構生物研究所研究助理
廖竣
臺灣大學漁業科學研究所碩士
林威任
清華大學生物資訊與結構生物研究所研究助理
陳若尹
清華大學生命科學系大五生
曾晴賢
清華大學生命科學系教授