文/汪良奇
.png) 壹、孢粉化石打哪來
壹、孢粉化石打哪來
孢粉是植物的孢子與花粉的總稱,其存在於植物的世代交替間,微小肉眼難以窺視,需要顯微鏡的幫助才能一窺究竟。對植物而言,孢粉是讓他們稱霸陸地的利器,也是世代交替中重要的一環。
蕨類成熟孢子體減數分裂後形成孢子並從孢子囊飛散而出。孢子在潮濕的環境會萌發成為配子體並分化出藏精器和藏卵器,產生的雌雄配子在潮濕環境時釋出,並完成交配成為新的孢子體。種子植物則產生花粉,花粉就是種子植物的雄配子體,藉由風、昆蟲甚至水的運送到達雌配子體來完成授粉。對生物而言,傳宗接代可是一等一的大事。植物為了增加成功交配(授粉)的機率,因此往往在生殖季(花季)時拼了命產生數以萬計的孢粉(圖1),以增加成功機率。舉例來說木本的垂枝樺(
Betula pendula)每個花藥大約能產生11160顆花粉、草本的裸麥(
Secale cereale)每個花藥的花粉產量更高達22360顆。可以想像當花季來臨時,空中除了飄散著花香外,更充滿著微細的花粉。然而,幸運兒往往是萬中取一,無緣完成他生物學上任務的孢粉就如同落塵般墜落凡塵,如果剛好掉到湖泊或沼澤裡,就會成為萬古不化的孢粉化石。
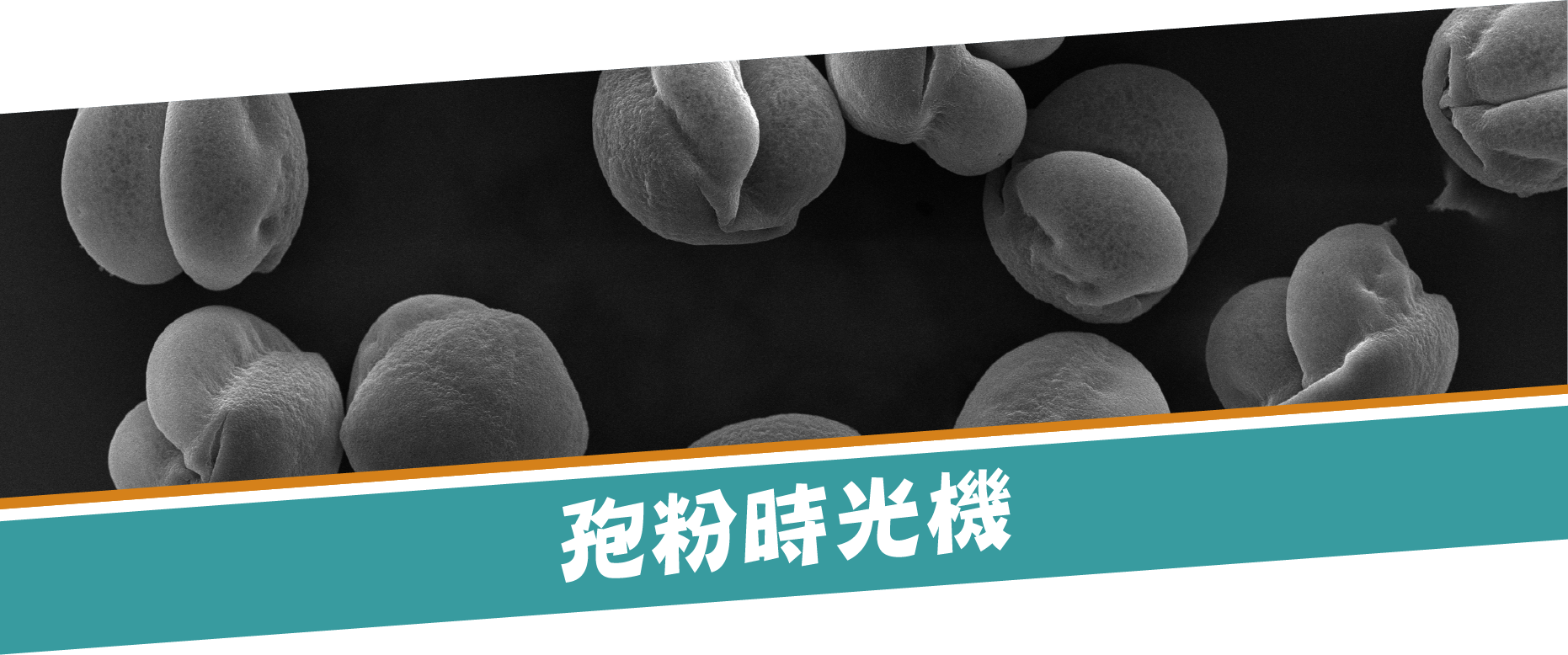
 圖1. 雲杉花粉。針葉樹多為風媒花,花粉產量極高。
圖1. 雲杉花粉。針葉樹多為風媒花,花粉產量極高。
貳、孢粉怎樣採集
陸地上的岩石風化後會變成碎屑,然後在低窪處堆積。沈積的過程中孢粉混合其中形成沉積物。湖泊是堆積與保存沉積物的良好環境,因此湖泊沉積物裡通常孢粉保存良好。另外生長在沼澤裡的苔蘚也能有效捕捉與保存沈降下來的花粉,也因此在富含有機質泥炭內也會保存大量的孢粉。利用各式沉積物岩心取樣器進行鑽取或在露頭剖面採樣,便能取得連續且不受擾動的沈積物。
取得沉積物後,下一步就是要將孢粉提取出來。孢粉的外壁是由孢粉質所組成,是一種非常惰性且能抵抗化學物質的反應。利用這種化學的特性,將沉積物泡到高濃度的鹽酸、氫氟酸裡便能把沉積物內的礦物如碳酸岩(如碳酸鈣)與矽酸岩(如石英、長石)融化移除,進而將沉積物內的孢粉濃縮。除了用暴力的化學手段外,也可以利用溫和的物理原理將孢粉濃縮。孢粉質的比重約為1.88,將沈積物置入比重大於2的無毒重液充分混合攪拌後靜置,比重較輕的孢粉便會浮到重液上層並與比重較重的礦物分離,進而達到濃縮的效果。為了讓孢粉在顯微鏡觀察時花粉外壁的形態特徵有較好的對比,因此處理過程中還會加入1:9的硫酸與醋酸混合液來將花粉外壁染色。大部分化石孢粉大小小於100微米,因此處理好的濃縮孢粉必須要先利用甘油膠製成永久玻片,才能在光學顯微鏡下進行鑑定。
參、孢粉長怎樣
孢粉是植物演化上重要的生殖器官,因此不同種類的孢粉形態有顯著的差異。第四紀(近258萬年)的化石孢粉鑑定屬於自然分類系統,也就是依據現生花粉形態來進行鑑定後,以現生植物命名。因此藉由現生孢粉形態資料庫的蒐集與建立,便能對第四紀的孢粉進行鑑定。早於第四紀的化石孢粉鑑定與命名,由於其花粉形態與現代植物已有明顯差別,難以直接置入現有的自然分類系統,因此命名系統上則使用半自然分類系統或形態分類法。
蕨類孢子形態較為單一,蕨類孢子母細胞減數分裂後,形成4分體,依四分體交界處(近心極)就會發育為萌芽溝,4分體分裂為單孢子依萌芽溝的形狀可分為單溝孢子與三溝孢子兩大類(圖2、圖3)。
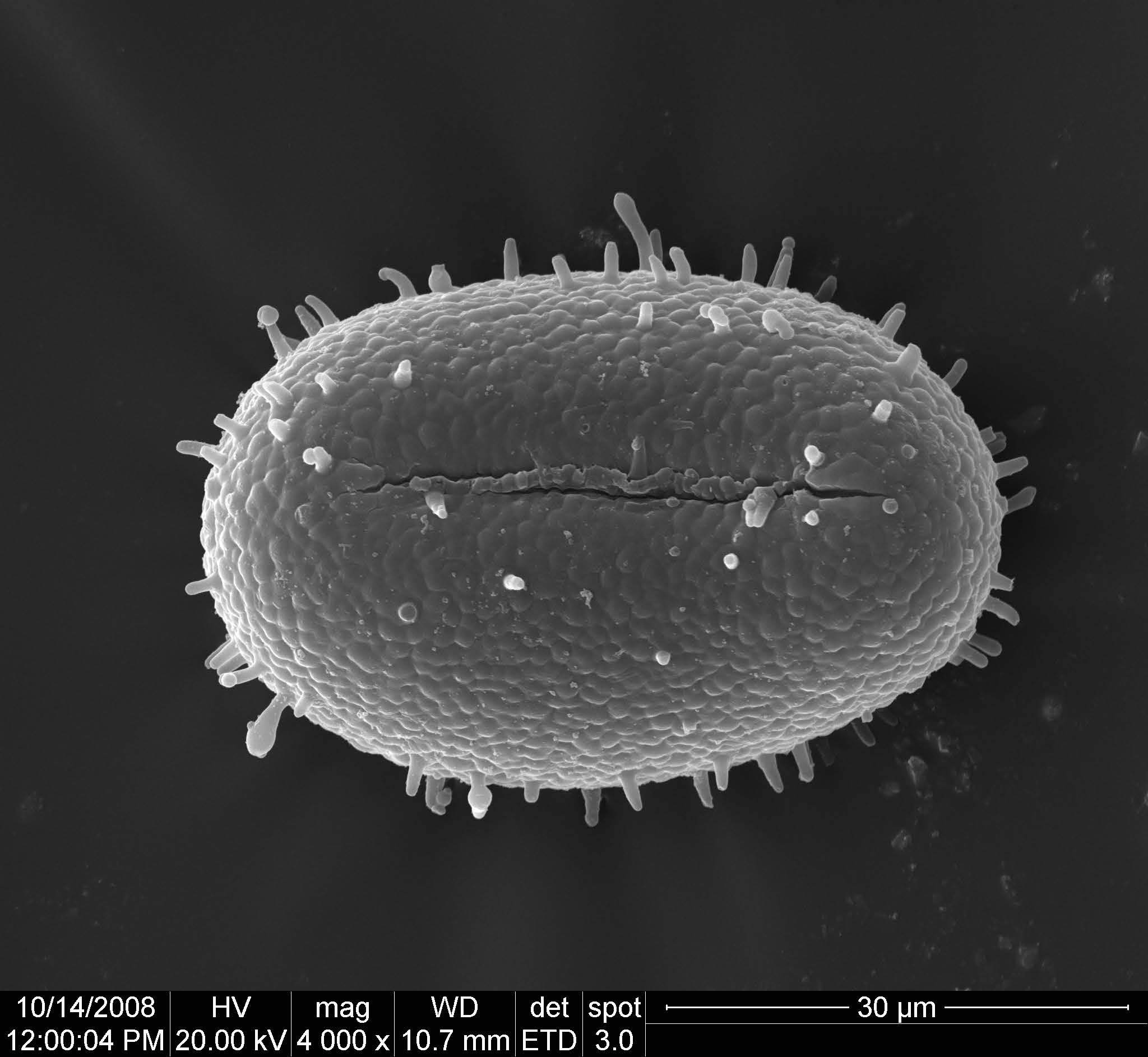 圖2. 蕨類單溝孢子。
圖2. 蕨類單溝孢子。
 圖3. 蕨類三溝孢子。
圖3. 蕨類三溝孢子。
花粉的形態則十分多變且複雜,花粉母細胞減數分裂後,所形成的四分體大部分在花粉成熟後會分開成為單花粉。但在某些科或屬的植物花粉會聚合合成複合花粉。如杜鵑科的4合花粉(圖4)與相思樹屬的16合花粉(圖5)。蘭科花粉則是由數以千計的花粉粒聚合成花粉塊。
 圖4. 杜鵑花粉,屬於4合花粉。
圖4. 杜鵑花粉,屬於4合花粉。
 圖5. 相思樹屬化石花粉,屬於16合花粉。
圖5. 相思樹屬化石花粉,屬於16合花粉。
由於花粉是立體的,在光學顯微鏡觀察下會有不同的面,因此在形態描述上需要定義不同的視角。花粉跟地球一樣有兩極與赤道,花粉的兩極是依據在4分體的位置所定義,朝內的面稱為近極面,朝外的面則稱為遠極面,兩個面的中心點則近極點與遠極點,兩個極點連接的線稱為極軸。與極軸垂直的平面則稱為赤道面。花粉萌芽孔發育位置通常座落在赤道面上,因此在顯微鏡觀察時,極面可以觀察到所有萌芽孔,而赤道面可以觀察到清楚的溝孔構造(圖6)。
 圖6. 黃苑花粉,左右兩顆為極面觀,中間為赤道面觀。
圖6. 黃苑花粉,左右兩顆為極面觀,中間為赤道面觀。
在了解不同視角的定義後,就可以量測花粉極軸與赤道軸,並計算其比值,藉此定義花粉的形狀。如比值小於0.5稱超扁球形;等於1稱圓球形而大於2稱超長球形。除了形狀,孢粉的大小也是描述的重點,如大於200微米稱為巨型;25-50微米稱中型(大部分化石花粉大小);小於10微米稱微型。
萌芽孔為花粉管伸出的位置,為花粉外壁上的開口或變薄處。萌芽孔的形狀為長條狀稱之為溝,形狀短或圓的萌芽孔稱之為孔。兩種類型的複合體稱為溝孔。結合花粉萌芽孔的類型、數量與位置便是花粉形態描述的重點(圖7)。花粉表面紋飾主要由花粉外壁形態所決定,其類型也是花粉種類判定的重要資訊。花粉外壁縱向剖面如同房子一般,可分為底層(地板)、柱狀層(柱子)與覆蓋層(屋頂)。覆蓋層上的突起或缺失、柱狀層的形狀與分佈,都會影響紋飾類型(圖8)。
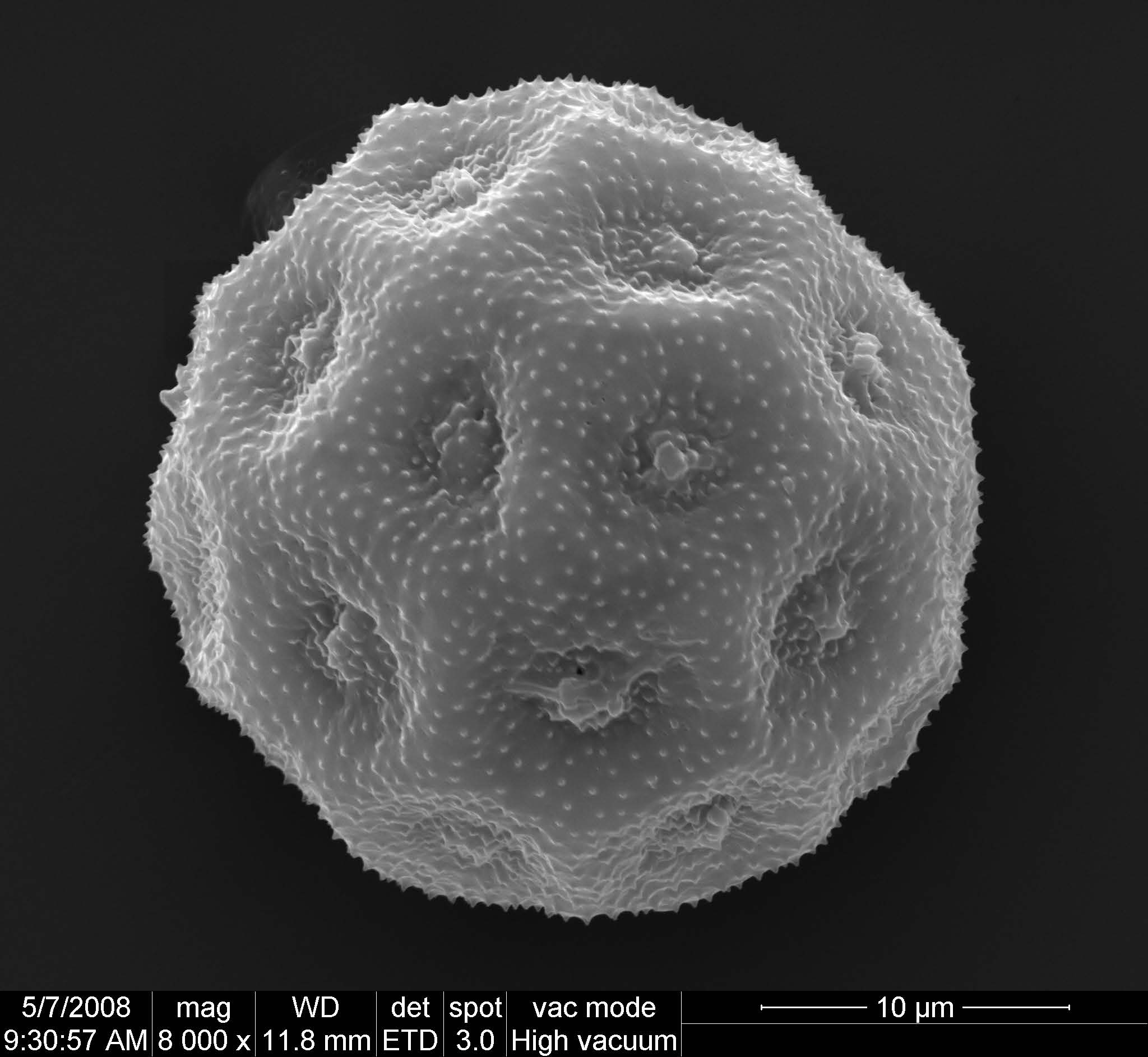 圖7. 澤瀉花粉,屬於多孔花粉。
圖7. 澤瀉花粉,屬於多孔花粉。
 圖8. 黃花杏菜花粉極面觀,有明顯條紋狀紋飾。
圖8. 黃花杏菜花粉極面觀,有明顯條紋狀紋飾。
結合上述的形態特徵便能進行花粉形態描述,舉例來說,冬青屬花粉(圖9)為中型3溝孔花粉。赤道面觀近長球形,溝寬且長,延伸到兩極。溝內有顆粒狀飾,溝間區則為孢芽狀紋飾。在良好的花粉描述下,不需要看到圖片,也能精準想像花粉形態。
 圖9. 冬青屬化石花粉。
圖9. 冬青屬化石花粉。
肆、孢粉如何重建古環境
化石孢粉濃縮與完成種類鑑定後,便可開始進行花粉的計數分析。一般來說每個沉積物樣本會計數300個花粉粒。為何是計數300顆花粉而不是計數到1000顆或更多花粉?這是因為在統計上,單個樣本計數到300個花粉後,主要種屬間的百分比差異變化小於1%。因此計數更多的花粉並不會影響優勢種類花粉在沉積物內百分比的比例。然而計數與鑑定花粉是非常費時費工的勞力活,因此每個樣本計數300顆花粉,是衡量所需耗費精力與資料誤差上所取得的平衡點。
接著要把計數的孢粉資料轉換為百分比,依需求可使用所有花粉計數的總和、部份特定花粉計數的總和(如木本植物花粉、陸生植物花粉)作為分母,將各類孢粉轉換成百分比後依植群歸屬分群排好,以深度或年代做縱軸繪製花粉圖譜。下一個步驟,則是進行花粉生物地層的劃分與判定。在連續的沉積物內,孢粉圖譜所呈現的組成與百分比變化代表過去周圍植群的組成與數量變化。因此當花粉組成有明顯改變時,通常代表明顯的環境改變。為了能客觀判斷花粉組成在哪個時間點為其組成轉變的邊界。可將花粉資料進行多變量分析中群聚分析,計算兩相鄰樣本間相似度差異,並依此繪製的樹狀圖來協助每個花粉生物帶邊界的判定。而孢粉圖譜的闡述與古環境的重建,便是基於每個花粉生物帶所進行。
以下則以位於宜蘭撤退池的全新世孢粉紀錄為例進行古環境重建(圖10)。在花粉生物帶RL1時(11000-8600 cal BP; cal BP為碳十四定年校準年代,以西元1950年為基準往回推算),草本花粉與溼地植物花粉含量高,木本花粉以高海拔針葉樹種鐵杉屬花粉最為優勢,可以想像當時針葉林帶向中海拔區域延伸,氣候寒冷,撤退池周圍遍佈溼地與草地。隨後在花粉帶RL2時(8600-6400 cal BP),中海拔闊葉林樹種昆欄樹屬與櫟屬取代鐵杉成為優勢樹種,反應中海拔闊葉林的擴張與十分溫暖的氣候。此時,中海拔闊葉森林取代先前在湖邊的草地與溼地。RL3(6400- 4500 cal BP)時,鐵杉再度逐漸增加,顯示氣候再度變得涼爽,較多的溼地植物花粉顯示當時撤退池溼地面積逐漸增加。最後RL4(4500 cal BP以來),在4500-2000 cal BP出現的沈積間斷,可能反應當時非常寒冷乾燥的氣候。2000 cal BP以來先驅樹種赤楊屬花粉成為最有優勢的種類,可能反應較多的擾動事件,如地震、颱風與山崩事件。溼地植物花粉的顯著增加,反應撤退池因泥炭堆積逐漸由湖泊轉變成為沼澤的過程。所重建的古環境可對古氣候進行推衍。在全新世早期(RL1)的寒冷氣候顯示東亞夏季風上處於微弱狀況、中全新的大暖期(RL2)除了反應台灣氣候最適宜的時期外,同時也代表東亞夏季風在本時期強度達到最強。與隨後4000 cal B開始的氣候變冷事件則與東亞夏季風減弱有關。與各種可能營力對比後,東亞夏季風強弱的變化趨勢大致與北緯30度日照強度變化有關,由此可知台灣全新世氣候變化趨勢大致與東亞夏季風強弱變化與太陽日曬變化趨勢有關。
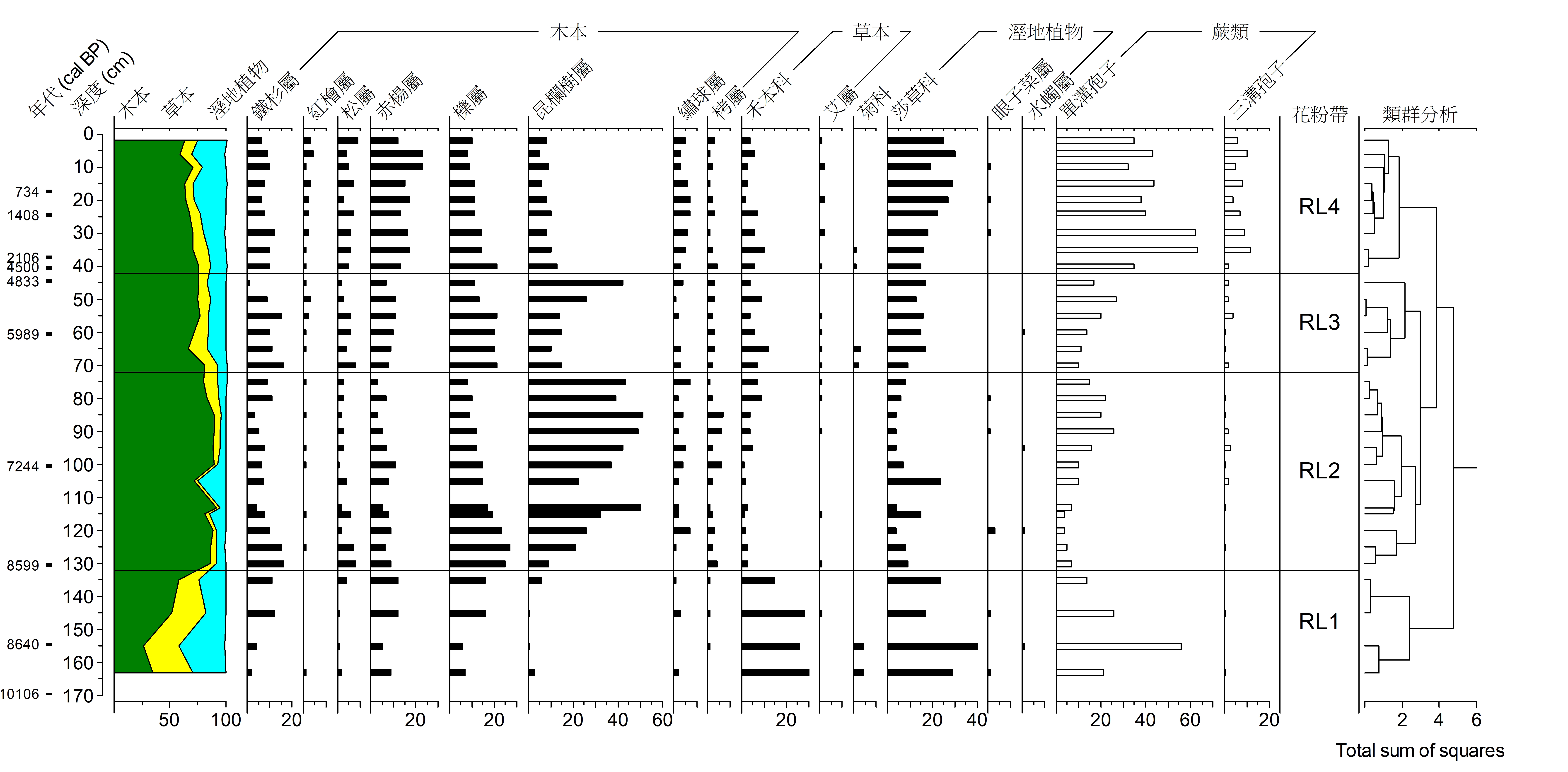 圖10. 撤退池花粉圖譜。
圖10. 撤退池花粉圖譜。
伍、孢粉看變遷
孢粉的變化紀錄著過去植群的變遷,而大災難的事件同時也會刻印在沉積物的孢粉紀錄內。隕石撞擊事件結束了恐龍盛行的白堊紀。在隕石撞擊之後,粉碎的隕石碎片,在全球地表堆積了一層富含銥元素的沈積層。分析該層的孢粉發現,花粉含量瞬間降到極低,為當時地表大部分陸生植物在這次撞擊事件當下幾乎全部滅絕的強烈證據。
近代人類活動與大量燃燒化石燃料,改變地球大氣組成,並造成了急速的全球暖化與生物多樣性的減少。然而人類對陸地上的植群的影響到底是從何時開始的。畢竟對生物的滅絕,在地球歷史上全球尺度的氣候變遷事件,如冰期與間冰期的轉換,也是十分重要的因素。以上次冰盛期為例,進入全新世後全球平均溫度大約上升了6度C左右。孢粉學家利用全球孢粉資料庫的資料,分析全球的各地共1181個化石孢粉紀錄的植群改變速率,來探討上次冰盛期以來氣候和人類活動對陸域生態系的影響。結果顯示全球陸域植物多樣性的變化主要自4600-2600 cal BP開始。晚全新世人類對陸域植物多樣性改變與上次冰期進入全新世時所造成的變化強度相當。人類活動對陸域生態系與多樣性的影響已達上次冰盛期與間冰期轉換的強度,而近百年的影響更是顯著加強。
另一個研究則探討人類移入對於島嶼植群的影響,該研究分析27個全不同大小島嶼的化石孢粉紀錄,嘗試釐清人類的遷入如何影響島嶼生態功能。在比較人類到達前後的植群組成改變,顯示人類到達後大約造成11倍的變化速度增加。這樣的結果也提供未來進行生態多樣性保育與生態復育前,需要考量人類遷入前後的原始生態系的差異。
在台灣化石孢粉的研究也提供許多古天災與人類活動的證據。在整合台灣東部與東北部4個湖沼沉積物的孢粉紀錄,發現在小冰期早期台灣東部降雨較多,但在小冰期中期後在東北部則有降雨增加的現象。這不對稱的區域降雨事件,可能紀錄了颱風路徑的改變。而颱風的路徑在全球氣候較冷時期時,由於颱風生成地區會偏向西太平洋,因此路徑會直接向西,並造成較多颱風通過台灣。反聖嬰現象盛行時,生成颱風的地點會稍微向西北移動。而在沉積物中觀察到東部與東北部降雨的差異,推測是源於小冰期初期與晚期反聖嬰現象頻率不同所致。根據孢粉所提供的資料推測在未來全球暖化的趨勢下,通過台灣的颱風可能變少,因此在台灣水資源管理上必須提前進行應變。